The role of morphology and microscopy in coccidian research in this genomic/proteomic age
David J. P. Ferguson
Nuffield Department of Pathology, Oxford University,
John Radcliffe Hospital, Oxford, OX3 9DU, UK
The history of the Coccidia is intimately involved with microscopy. The parasites are below the resolution of the human eye (with the exception of giant tissue cysts of certain Sarcocystis species) therefore microscopy is required for visualisation and identification of the parasite. The Coccidia belong to the phylum Apicomplexa, a widespread and successful group of protozoan parasites which includes many of medical and veterinary importance. The family Coccidia is characterised by a life cycle involving faecal/oral transmission with asexual and sexual development within the alimentary tract of the definitive host. To survive the rigors of the external environment the parasites have evolved a cystic stage (oocyst) that is released with the faeces and can survive in the external environment. The appearance and size of these oocysts are characteristic of a given species and microscopy of faecal samples has been used for many years to diagnose infections and to identify the species involved. This single host species life cycle is typical of those parasites of the genus Eimeria responsible for coccidiosis in chickens. However, a sub-group of the Coccidia (including the genera Toxoplasma, Neospora, Hammondia, Besnotia and Sarcocystis) has evolved a more elaborate life cycle involving intermediate hosts where tissue cysts are formed in the muscle or brain. In these cyst forming Coccidia, infection can be transmitted to the definitive host when ingested with the tissue of the intermediate host. There are obvious advantages for this complex life cycle for coccidian parasites of carnivores.
The vital role of microscopy, especially electron microscopy, can be inferred from the name of the phylum the Apicomplexa. This name is derived from the ultrastructural identification of a characteristic group of organelles at the apical end of the infectious stages. Through the 1970s and 80s straightforward electron microscopy added greatly to knowledge and understanding of the Coccidia and the developmental processes involved in their life cycle. Unfortunately many of the papers were published before the modern electronic age and therefore are not readily accessible. More recently there has been a renaissance of both light and electron microscopy with the development of new molecular techniques. A major advance was the application of immunocytochemistry in which antibodies can be used to identify and localise specific molecules during the parasite life cycle. Now light and electron microscopy, in conjunction with modern genomic and proteomic techniques, plays a vital part in a multidisciplinary approach to improve our understanding of Coccidian parasites. A few areas where microscopy and immunocytochemistry have added to our knowledge of the Coccidia will be illustrated.
Apical complex
The specific group of organelles found in the anterior of the infectious stages
that give rise to the name Apicomplexa consist of three distinct structures; the
micronemes (small cigar-shaped structures), the rhoptries (bulbous structures
with ducts running to the anterior) and the dense granules (spherical structures)
that can also be found in other regions of the parasite (Fig 1a). In the
Coccidia, the infectious stages also possess an additional organelle, the conoid,
which is a truncated cone comprised of microtubules (Fig 1a). The relative and
absolute numbers of the various organelles can vary between species and even
between the infectious stages in a single species. For example the merozoites of
Eimeria tenella have large numbers of micronemes but few dense granules in
contrast to the tachyzoites Toxoplasma gondii that have numerous dense granules
but few micronemes. Within the Coccidia, this group of organelles are believed
to play a co-ordinated role in the infection of new host cells. The contents of
the micronemes are released first and are involved identification and adherence
to suitable host cells. The content of the rhoptries are released during the
invasion process and some proteins are incorporated into the parasitophorous
vacuole. Finally the dense granules are secreted into and are believed to modify
the parasitophorous vacuole to facilitate parasite development (reviewed by
Carruthers 2002, Mercier et al 2005). Using molecular techniques, numerous new
proteins are continually being identified (Zhou et al 2005, Bradley et al 2005)
and by using immuno-light and electron microscopy it has been possible to
identify the location of certain of these molecules to one or other of the
apical organelles. This can assist in understanding their biological function.
By immuno-light microscopy, molecules can be localised to the apical region (Fig
1b) and using immuno-electron microscopy it is possible to identify the exact
organelle they are located in (Fig 1c). It is possible to localise unknown
apical proteins to the micronemes (MIC proteins), the rhoptries (ROP proteins)
or the dense granules (GRA proteins). It is also possible using double or triple
labelling, to show that different proteins are located within the same or
different organelles. For example it can be confirmed that GRA2, 4 and 6 are
located within the same dense granule (Fig 1d) and it is also possible to
differentiate between organelles by their protein content with rhoptries
labelled with anti-ROP2 and dense granules by anti-NTPase, a dense granule
protein (Fig 1e). In elegant studies using T. gondii, which is most amenable to
molecular manipulation, it has been possible to transfect parasites with
fluorescent (GFP) proteins and identify the factors which specifically targets
proteins to each of the various organelles (Gubbels and Striepen, 2004).
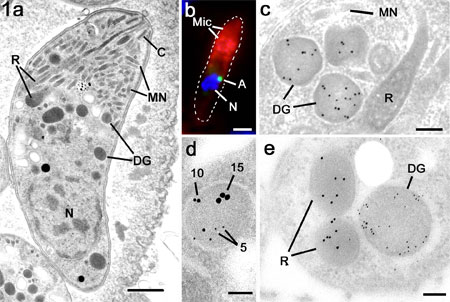
Figure 1a. Electron
micrograph of a bradyzoite of T. gondii showing the characteristic compliment
of apical organelles consisting of the conoid (C), micronemes (MN), rhoptries
(R) and dense granules. N – nucleus. Bar is 1mm.
b. Immuno-fluorescent image of a double labelled merozoite
of E. tenella showing labelling of the anterior with anti-MIC2 (Mic; red) and
the small peri-nuclear apicoplast labelled with anti-enoyl reductase (A; green).
Nucleus labelled with DAPI (N; blue). Bar is 1mm.
c. Immuno-electron micrograph of a tachyzoite of T. gondii
labelled with anti-GRA6 showing the gold particles are limited to the dense
granules (DG) while the micronemes (MN) and rhoptries (R) and negative. Bar
is 100 nm.
d. A triple labelled section using anti-GRA2 (5 nm gold), anti-GRA4
(10 nm gold) and anti-GRA6 (15 nm gold) showing a dense granules labelled with
all three markers. Bar is 100 nm.
e. Cross section through the anterior of a merozoite of T.
gondii double labelled with anti-ROP2 (10 nm gold) and anti- NTPase (5 nm gold)
showing the specific labelling of the rhoptries (R) and dense granules (DG).
Bar is 100nm
(Copyright for all images is retained by D J P Ferguson, Oxford University)
Apicoplast
More recently, the apicomplexan parasites have also
been found to possess a second unique feature. They contain a non-photosynthetic
plastid, which has been termed the apicoplast (reviewed by Wilson 2002, Waller
and McFadden 2005). This residual plastid is believed to be derived from an
ancient secondary endosymbiotic acquisition from a red alga (Wilson 2002). The
plastid genome was first identified in Plasmodium sp. but has subsequently been
shown to be present in all members of the Apicomplexa with the exception of
Cryptosporidium sp. (Xu et al 2004). It has been shown that the apicoplast is
essential for parasite survival and parasites without an apicoplast are non-viable.
In the absence of a plastid, parasites displayed an unusual delayed death phenotype
(He et al 2001). That is, the parasites without a plastid could continue to
divide within the host cell vacuole as long as one plastid containing organism
was present. Indeed they were able to escape and invade new host cells but were
unable to undergo proliferation within the new host cell resulting in death
of the parasite (He et al, 2001). These findings are extremely exciting since
the prokaryotic nature of this organelle could represent new and unique targets
for drug intervention. The role of the apicoplast is still incompletely understood
but it is known to be involved in type II fatty acid biosynthesis and isoprenoid
biosynthesis (reviewed Wilson 2002, Waller and McFadden 2005). By electron microscopy
it was shown that this genome was located within an organelle characterised
by being limited by multiple membranes (McFadden et al 1996, Kohler et al 1997)
(Fig 2c). This organelle was first identified by electron microscopy in the
1960s (Sheffield and Melton 1968) although its function remained unknown. The
exact number of membranes is still controversial (Kohler 2005) but the presence
of multiple (3/4) membranes allows identification of the organelle and differentiates
it from the other genome containing organelles (the nucleus and the mitochondrion)
that are limited by two membranes (Fig 2c, e). It has been found that many of
the plastid specific proteins are encoded by nuclear genes (lateral gene transfer)
and these proteins have to be targeted back to the apicoplast using a specific
bi-partite targeting sequence. Initial morphological studies of the apicoplast
examined, in vitro, asexual development of parasites transfected with the green
fluorescent protein targeted to the apicoplast (Striepen et al 2000, Vaishnava
et al 2005). More recently, in vivo studies using antibodies to an apicoplast
specific protein (enoyl reductase) have examined the changes in the apicoplast
during both asexual and sexual development in T. gondii (Ferguson et al 2005)
and E. tenella. In the infectious (sporozoite, merozoite, tachyzoite
and bradyzoite) stages the apicoplast is normally present as a single small
spherical structure adjacent to the nucleus (Fig 1b, 2a) although multiple plastids
have been observed in the merozoites of E. tenella (Fig 2b). Asexual
proliferation (schizogony) results in the simultaneous formation of numerous
daughters. To ensure their viability, each merozoite must receive a nucleus
and at least one apicoplast and mitochondrion. This requires a tightly co-ordinated
process of nuclear, apicoplast and mitochondrial division and segregation. The
possible mechanisms are still under investigation. There are conflicting observations
with certain studies showing a close relationship between nuclear and apicoplast
division (Striepen et al 2000, Vaishnava et al 2005) while others show evidence
that nuclear and apicoplast division are independent events (Ferguson et al
2005) (Fig 2d). However, organelle segregation to the daughters may involve
the nuclear pole/centriolar complex in all cases. When sexual development was
examined, there was found to be little change in the apicoplast during microgametogony
and the microgametes lacked an apicoplast. In contrast there is a marked increase
in the size and activity of the apicoplast during macrogametogony perhaps reflecting
the increased metabolic activity (Fig 2f). This means that there will be maternal
inheritance of the apicoplast with the sporozoites, formed within the oocyst,
receiving their apicoplast from the macrogamete (Ferguson et al 2005).
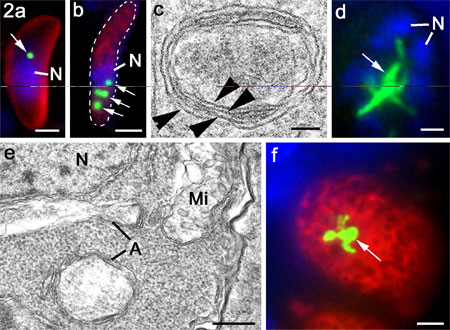
Figure 2 a and b.
Tachyzoite of T. gondii (a) and merozoite of E. tenella (b) labelled with anti-enoyl
reductase showing the peri-nuclear location of the single or multiple apicoplasts
(arrows; green). Bar is 1mm.
c. High power electron micrograph of an apicoplast in E. tenella
showing the four limiting membranes (arrowheads). Bar is 100 nm.
d. Immuno-fluorescent image of multinucleate schizont of T.
gondii in the cat intestine showing a large elongated and branched apicoplast
(arrows). Bar is 1mm.
e. Electron micrograph of part of a schizont similar to that
in d showing the enlarged multimembraned apicoplast (A) and the nucleus (N)
and mitochondrion Mi) limited by two membranes. Bar is 100 nm
f. Immuno-fluorescent image of macrogamete of T. gondii in
the cat intestine showing a large elongated and branched apicoplast (arrows).
Bar is 1mm.
(Copyright for all images is retained by D J P Ferguson, Oxford University)
Asexual development
Asexual development within the Coccidia is often
considered as a single process, which has been termed schizogony, only varying
in the number of daughters formed. However when the process was examined in
different species by electron microscopy, subtle differences could be identified
showing at least four variations in the process exhibited by different parasites.
There is classical schizogony which is undergone by the vast majority of coccidian
species including the genera Eimeria. It can be summarised as involving
a proliferative phase in which there are repeated cycles of DNA replication
and nuclear division giving rise to a multinucleate stage followed by a differentiation
phase where daughter formation is initiated at the parasite surface and the
nuclei and other organelles segregate to the daughters as they bud into the
parasitophorous vacuole (Fig 3). A variation of this process is undergone by
the coccidian stages of T. gondii in the cat gut where there is an identical
proliferative phase but during the differentiation phase daughter formation
is initiated and completed within the mother cell cytoplasm (Ferguson et al
1974, 2005) and this process has been termed endopolygeny (Fig 3). A third variation
is that undergone by certain asexual stages of Sarcocystis spp. where there
are repeated cycles of DNA replication but with no nuclear division. In this
case the initiation of multiple daughter formation occurs within the mother
cell cytoplasm and coincides with fragmentation of the polyploid nucleus into
a number of haploid nuclei each of which enters a developing daughter in a similar
manner to that seen in endopolygeny (Speer and Dubey 1999, 2001, Vaishnava et
al 2005). The fourth variation is the one that has been most extensively studied
since it is that undergone by the exo-enteric (tachyzoites and bradyzoites)
forms of T. gondii and has been termed endodyogeny. In this case only two daughters
are formed with the initiation of daughter formation within the mother cell
cytoplasm occurring before completion of nuclear division (Fig 3). The process
of daughter formation is similar to that observed during endopolygeny but only
two daughters are formed (Sheffield and Melton1968, Striepen et al 2000). These
three variations from classical schizogony are the exception and it may be significant
that they are found within the sub-group of cyst forming Coccidia, which have
had to adapt to development in intermediate hosts. Although these subtle variations
can only be identified by electron microscopy, an appreciation of these differences
can help to reconcile certain apparent conflicting results in relation to the
division and segregation of the nuclei and apicoplasts during daughter formation
in T. gondii, Sarcocystis sp., and E. tenella (Striepen et al
2000, Vaishnava et al 2005, Ferguson et al 2005).
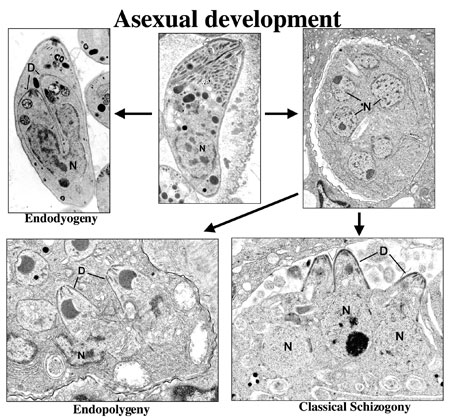
Figure 3a. A montage of asexual
development illustrating certain of the ultrastructural differences associate
with the process of endodyogeny, endopolygeny and classical schizogony within
the Coccidia.
(Copyright for all images is retained by D J P Ferguson, Oxford University)
Sexual development
Microgamete formation
Within the Coccidia, microgametogony results in the
formation of relatively low numbers of microgametes. The relative number of
microgametocytes varies between species. In the case of T. gondii, where there
are relatively few microgametocytes to the number of macrogametes, it has been
questioned if there are sufficient microgametes to ensure that efficient fertilisation
can take place (Ferguson 2003). By electron microscopy it is interesting to
note just how similar microgametes are to mammalian sperm. They consist of a
compact, electron dense nucleus, a mitochondrion and are powered by two large
flagella (Fig 4a, b).
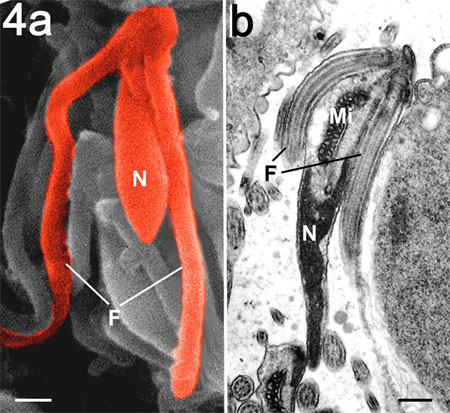
Figure 4. Scanning (a) and transmission electron microscopy
(b) of the mature microgamete of T. gondii showing the nucleus
(N), mitochondrion (Mi) and two flagella (F). Bar is 100 nm.
(Copyright for all images is retained by D J P Ferguson, Oxford University)
Macrogamete and oocyst formation
The survival of this group of parasites depends on
the ability of the oocysts formed from the macrogametes to survive for extended
periods in the external environment. Not only that, but the oocyst have to undergo
sporulation to form eight infectious sporozoites, which is the only example
of extra-cellular development by coccidian parasites. This places two essential
requirements on the developing macrogamete. The first is that it must acquire
and store all the ingredients necessary to allow sporulation while cut off from
additional nutrients in the hostile external environment. Thus the macrogamete
contains large amounts of storage material in the form of polysaccharide granules
and lipid droplets (Fig 5a). The second is for the macrogametocyte to synthesise
and store the material required to form the oocyst wall. The oocyst wall is
a complex structure consisting of a loose outer veil, which is lost during excretion,
and a wall consisting of two distinct layers, a thick electron dense outer layer
and a thinner more electron lucent inner layer (Fig 5g). The structure is pretty
consistent across the Coccidia (Ferguson et al, 1975, 1977, 2003). The report
of a thin (10 nm thick) outer layer and a thicker (90nm) inner layer observed
for oocysts of E. tenella isolated from faeces (Stotish et al
1978) may represent an example of where part of the wall has been lost during
processing. Any disruption in the process of oocyst wall formation would have
serious implications for parasite transmission. To understand this process,
genes for certain components of the oocyst wall have been identified (Belli
et al 2002a, b, 2003, Wallach, 2002). Using antibodies to these and to other
proteins it has been possible to start to dissect the changes occurring during
macrogamete maturation and oocyst wall formation by combining electron microscopy
and immunocytochemistry (Mouafo et al. 2002, Ferguson et al, 2003). An accidental
and unexpected finding was that an antibody to the specific apple domains of
the MIC4 protein of T. gondii (TgMIC4, Brecht et al 2001) cross reacted with
a subset of granules in the early macrogametocyte of T. gondii (Ferguson et
al, 2000) but also with small granules in the early macrogametocytes of E.
maxima (Ferguson et al, 2003) and E. tenella. In all three
cases, the contents of these granules were secreted into the parasitophorous
vacuole and appeared to associate with the formation of the loose outer veil
(Fig 6d). These previously unidentified granules have been termed the veil forming
bodies (VFB) (Ferguson et al 2003). It is interesting to speculate that the
veil contains a protein with apple domains and, since these domains are associated
with protein-protein or protein-carbohydrate interactions, they could be involved
in microgamete recognition. The second type of granule is the large electron
dense membrane bound spherical structures, which have been termed the wall forming
bodies type 1 (WFB1) (Fig 5a, g). In addition, there is a third structure consisting
of irregular shaped electron dense deposits located with the rough endoplasmic
reticulum that have been identified as the wall forming bodies type 2 (WFB2)
(Fig 5a, g). Using immuno-light and electron microscopy is was possible to identify
and follow the changes in these structures during oocyst wall formation. Antibodies
to affinity purified gametocyte antigens (anti-APGA, Belli et al 2002a, b) stained
the WFB1 strongly and the WFB2 weakly (Fig 5c) and it was possible to follow
the secretion of the WFB1 to form the outer layer of the oocyst wall (Fig 5e,
f). In contrast, the anti-gam56 (Belli et al 2002b) and anti-gam82 (Belli et
al, 2003a) antibodies only stained WFB2 (Fig 5d), the contents of which were
released to form the inner layer of the oocyst wall (Fig 5f). The question of
how material located in the endoplasmic reticulum could be secreted remained.
However, by immuno-EM, it was observed that, only after the release of the contents
of WFB1, the WFB2 material retained in the rER was transferred to the surface
via the Golgi body (Ferguson et al, 2003). This sophisticated control mechanism
allows independent and sequential secretion of the three components thus allowing
efficient formation of the stratified oocyst wall. These results were obtained
using antibodies to Eimeria maxima proteins but there is likely
to be close homology between the genes within the genus Eimeria and indeed
it was found that these antibodies cross reacted with the homologous proteins
of E. tenella and gave a similar staining pattern.
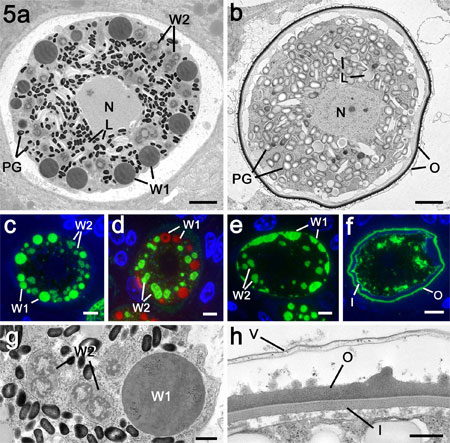
Figure 5a. Electron micrograph through a mature macrogamete of E. maxima
showing the central nucleus and the cytoplasm packed with numerous polysaccharide
granules (PG) and lipid droplet (L) in addition to the WFB1 (W1) and the WFB2
(W2). Bar is 1mm.
b. Sections through an early oocyst with a fully formed oocyst
wall. Note that while the polysaccharide granules (PG) and lipid droplets (L)
are still present but both types of WFB had disappeared. Bar is 1mm.
c. Immuno-fluorescent images through a mature macrogamete stained
with anti-APGA showing strong labelling of the WFB1 (W1) and lighter staining
of the WFB2 (W2). Bar is 1mm.
d. Similar organism to that in c stained with anti-gam56 showing
that only the WBF2 (W2) are stained. The WFB1 (W1) were visualised by Evans
blue staining (red). Bar is 1mm.
e. Early stage of oocyst wall formation showing secretion of
the WFB1 to initiate formation of the outer layer of the oocyst wall (W1). Stained
with anti-APGA. W2 – WFB2. Bar is 1mm.
f. Late stage in oocyst wall formation in which both the inner
(I) and outer (O) layers are formed. Stained with anti-APGA. Bar is 1mm.
g. Detail from the macrogamete in a showing the large membrane
bound WFB1 (W1) and the smaller irregular clumps of material representing the
WBF2 (W2) located within the rER. Bar is 0.5 mm.
h. Detail of the fully formed oocyst wall consisting of the
loosely attached outer veil (V) along with the electron dense outer layer (O)
and the thinner and less electron dense inner layer (I). Bar is 0.5 mm.
(Copyright for all images is retained by D J P Ferguson, Oxford University)
The process of fertilisation is yet to be observed in detail but it would appear
that oocyst wall formation occurs prior to release from the host cell (Fig 6b).
An extensive search failed to show microgametes associated with the macrogametes
even when located in adjacent cells (Fig 6a). However, a high proportion of
oocysts in the lumen were found to have attached microgametes (Fig 6c). These
were located beneath the veil adjacent to the outer layer of the wall (Fig 6d).
It is still not clear when and how the microgamete completes fertilisation.
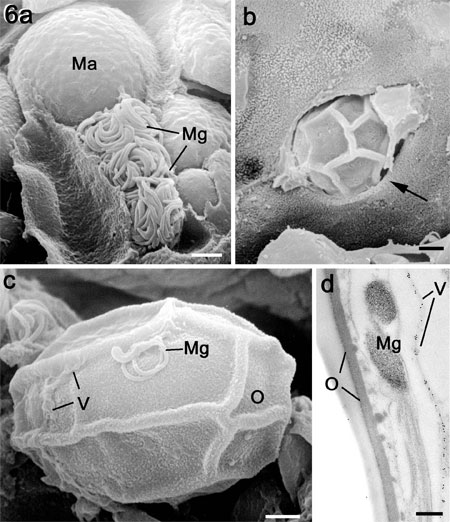
Figure 6a. Scanning electron micrograph (SEM) of a fracture through
a caecum infected with E. tenella showing a microgametocyte with numerous mature
microgametes (Mg) in a cell adjacent to a macrogamete (Ma). Bar is1 mm.
b. SEM of the luminal surface of a crypt showing an oocyst
(characterised by the surface folds) escaping from and epithelial cell (arrow).
Bar is1 mm.
c. SEM of an oocyst in the gut lumen showing partial loss of
the veil (V) and with a microgamete (Mg) adhering to the outer layer of the
oocyst wall (O). Bar is1 mm.
d. Transmission electron micrograph through the surface of
an oocyst showing the microgamete (Mg) located between the veil (labelled with
anti-TgMIC5) (V) and the outer layer (O) of the oocyst wall. Bar is 100nm.
(Copyright for all images is retained by D J P Ferguson, Oxford University)
This summarises a few examples where microscopy, combined with immunocytochemistry,
has helped in our understanding of the Coccidia. There are still many areas
where further research, using molecular techniques and microscopy, will provide
new insights and perhaps identify areas of weakness in this important group
of parasites.
This summarises a few examples where microscopy, combined with immunocytochemistry, has helped in our understanding of the Coccidia. There are still many areas where further research, using molecular techniques and microscopy, will provide new insights and perhaps identify areas of weakness in this important group of parasites.
Acknowledgements
The work described in this paper has only been possible
because of the active collaboration of a large number of people from the research
groups of Rima McLeod (Chicago University, USA), Craig Roberts (Strathclyde
University, UK), John Boothroyd (Stanford University), USA, Fiona Tomley and
Martin Shirley (Compton, UK), Sabina Belli and Nick Smith (University of Technology,
Sydney), Dominique Soldati (Geneva, Switzerland), J-F Dubremetz (Montpellier,
France) and M-F Cesbron-Delauw (La Tronche, France). DJPF was supported by an
equipment grant from the Wellcome Trust. The work on the structure of the macrogamete
and oocyst was supported by ABIC Ltd.
References
Belli SI, Lee M, Thebo P, Wallach MG, Schwartsburd
B, Smith NC, 2002a. Biochemical characterisation of the 56 and 82 kDa immunodominant
gametocyte antigens from Eimeria maxima. Int J Parasitol. 32, 805-16.
Belli SI, Wallach MG, Smith NC. 2003. Cloning and characterization of the 82
kDa tyrosine-rich sexual stage glycoprotein, GAM82, and its role in oocyst wall
formation in the apicomplexan parasite, Eimeria maxima. Gene 307, 201-12.
Belli SI, Witcombe D, Wallach MG, Smith NC,
2002b. Functional genomics of gam56: characterisation of the role of a 56 kilodalton
sexual stage antigen in oocyst wall formation in Eimeria maxima. Int J Parasitol.32,1727-37.
Bradley PJ, Ward C, Cheng SJ, Alexander DL, Coller S, Coombs GH, Dunn JD, Ferguson
DJP, Sanderson SJ, Wastling JM, Boothroyd JC. Proteomic analysis of rhoptry
organelles reveals many novel constituents for host-parasite interactions in
Toxoplasma gondii. J Biol. Chem. Jul 7; [Epub ahead of print]
Brecht S, Carruthers VB, Ferguson DJP, Giddings OK, Wang G, Jackle U, Sibley
LD, Soldati D. 2001. The Toxoplasma microneme protein MIC4 is an adhesin composed
of six conserved apple domains J. Biol. Chem. 276, 4119-27.
Carruthers VB. 2002. Host cell invasion by the opportunistic pathogen Toxoplasma
gondii. Acta Trop. 81:111-22.
Ferguson DJP. 2002 Toxoplasma gondii and sex: essential or optional extra. Trends
in Parasitology 18: 355-359.
Ferguson DJP, Belli SI, Smith NC, Wallach MG. 2003. The development of the macrogamete
and oocyst wall in Eimeria maxima: Immuno-light and electron microscopy. Inter
J Parasitol 33: 1329-1340, 2003.
Ferguson DJP, Birch-Andersen A, Hutchison WM, Siim JChr. 1977. Ultrastructural
studies on the endogenous development of Eimeria brunetti IV. Formation and
structure of the oocyst wall. Acta Path Microbiol Scand, Sect B 85: 201-211.
Ferguson DJP, Brecht S, Soldati D. 2000. The microneme protein MIC4, or a MIC4-like
protein, is expressed within the macrogamete and associated with oocyst wall
formation in Toxoplasma gondii. Int. J Parasitol. 30,203-09.
Ferguson DJP, Hutchison WM, Dunachie JF and Siim J Chr. 1974. Ultrastructural
study of early stages of asexual multiplication and microgametogony of Toxoplasma
gondii in the small intestine of the cat. Acta Path Microbiol Scand, Sect B
82: 167-181.
Ferguson DJP, Hutchison WM, Siim, JChr. 1975. The ultrastructural development
of the macrogamete and formation of the oocyst wall of Toxoplasma gondii. Acta
Path. Microbiol. Scand., Sect. B 83, 491-505.
Gubbels MJ, Striepen B. 2004. Studying the cell biology of apicomplexan parasites
using fluorescent proteins. Microsc Microanal.10:568-79.
He, CY, Shaw MK, Pletcher CH, Striepen B, Tilney LG, Roos DS. 2001. A plastid
segregation defect in the protozoan parasite Toxoplasma gondii. EMBO J. 20:330-9.
Kohler S. 2005. Multi-membrane-bound structures of Apicomplexa: I. the architecture
of the Toxoplasma gondii apicoplast. Parasitol Res. 96:258-72.
Kohler S, Delwiche CF, Denny PW, Tilney LG, Webster P, Wilson PRG, Palmer JD,
Roos DS. 1997. A plastid of probable green algal origin in Apicomplexan parasites.
Science 275:1485-9.
McFadden, GI, Reith ME, Munholland J, Lang-Unnasch N. 1996. Plastid in human
parasites. Nature 381:482.
Mercier C, Adjogble KD, Daubener W, Delauw MF. 2005. Dense granules: Are they
key organelles to help understand the parasitophorous vacuole of all apicomplexa
parasites? Int J Parasitol. 35:829-49.
Mouafo AN, Weck-Heimann A, Dubremetz J-F, Entzeroth R. 2002. Monoclonal antibodies
specific for the two types of wall-forming bodies of Eimeria tenella macrogametes
(Coccidia, Apicomplexa). Parasitol. Res. 88, 217-24.
Sheffield, HG., Melton ML. 1968. The fine structure and reproduction of Toxoplasma
gondii. J. Parasitol. 54: 209-226.
Speer CA, Dubey JP. 1999. Ultrastructure of shizonts and merozoites of Sarcocystis
falcatula in the lungs of budgerigars (Melopsittacus undulatus). J Parasitol.
85:630-7.
Speer CA, Dubey JP. 2001. Ultrastructure of schizonts and merozoites of Sarcocystis
neurona. Vet Parasitol. 95:263-71.
Stotish RL, Wang CC, Meyenhofer M. 1978. Structure and composition of the oocyst
wall of Eimeria tenella. J Parasitol. 64:1074-81.
Striepen B, Crawford MJ, Shaw MK, Tilney LG, Seeber F, Roos DS. 2000. The plastid
of Toxoplasma gondii is divided by association with the centrosomes. J. Cell
Biol. 151:1423-34.
Vaishnava1 S, Morrison DP, Rajshekhar Y. Gaj RY, Murray JM, Entzeroth R, Howe
DK, Striepen B. 2005. Plastid segregation and cell division in the apicomplexan
parasite Sarcocystis neurona. J Cell Sci. (in press)
Wallach, M. 2002. The development of CoxAbic® a novel vaccine against coccidiosis.
World Poultry 18, 2-4
Waller RF, McFadden GI.2005. The apicoplast: a review of the derived plastid
of apicomplexan parasites. Curr Issues Mol Biol. 7:57-79.
Wilson, RJM. 2002. Progress with parasite plastids. J. Mol. Biol. 319:257-274.
Xu P, Widmer G, Wang Y, Ozaki LS, Alves JM, Serrano MG, Puiu D, Manque P, Akiyoshi
D, Mackey AJ, Pearson WR, Dear PH, Bankier AT, Peterson DL, Abrahamsen MS, Kapur
V, Tzipori S, Buck GA. 2004. The genome of Cryptosporidium hominis. Nature.
431:1107-12.
Zhou XW, Kafsack BF, Cole RN, Beckett P, Shen RF, Carruthers VB. 2005. The opportunistic
pathogen Toxoplasma gondii deploys a diverse legion of invasion and survival
proteins. J Biol Chem. 2005 Jul 7; [Epub ahead of print].